Lípidos

Los lípidos más simples son los ácidos grasos, y tienen la fórmula general
R—COOH, donde R representa una cadena de hidrocarburo. Los ácidos grasos son
componentes de muchos tipos más complejos de lípidos, incluyendo los triglicéridos o
triacilgliceroles, los glicerofosfolípidos y los esfingolípidos. Los lípidos que contienen
grupos fosfato se llaman fosfolípidos y los que tienen grupos esfingosina y carbohidrato
a la vez se llaman glicoesfingolípidos. Los esteroides, las vitaminas lipídicas y los terpenos
se relacionan con la molécula de isopreno, de cinco carbonos, y por consiguiente se
llaman isoprenoides. El nombre terpenos se ha aplicado a todos los isoprenoides, pero
en general se restringe a los que existen en las plantas.
Los lípidos tienen funciones biológicas diversas, al igual que estructuras diversas.
Las membranas biológicas contienen una variedad de lípidos anfipáticos, incluyendo
los glicerofosfolípidos y los esfingolípidos. En algunos organismos, la función de los
triacilgliceroles (grasas y aceites) es de moléculas intracelulares de almacenamiento de
energía metabólica. También las grasas suministran aislamiento térmico y amortiguamiento
a los animales. Las ceras en las paredes celulares, en los exoesqueletos y en la
piel protegen a las superficies de algunos organismos. Algunos lípidos tienen funciones
muy especializadas. Por ejemplo, las hormonas esteroides regulan e integran una variedad
de actividades metabólicas en los animales, y los icosanoides participan en la regulación
de la presión sanguínea, en la temperatura corporal y en la contracción de los
músculos lisos en los mamíferos. Los gangliósidos y otros glicoesfingolípidos están en
la superficie celular y pueden participar en el reconocimiento celular.
R—COOH, donde R representa una cadena de hidrocarburo. Los ácidos grasos son
componentes de muchos tipos más complejos de lípidos, incluyendo los triglicéridos o
triacilgliceroles, los glicerofosfolípidos y los esfingolípidos. Los lípidos que contienen
grupos fosfato se llaman fosfolípidos y los que tienen grupos esfingosina y carbohidrato
a la vez se llaman glicoesfingolípidos. Los esteroides, las vitaminas lipídicas y los terpenos
se relacionan con la molécula de isopreno, de cinco carbonos, y por consiguiente se
llaman isoprenoides. El nombre terpenos se ha aplicado a todos los isoprenoides, pero
en general se restringe a los que existen en las plantas.
Los lípidos tienen funciones biológicas diversas, al igual que estructuras diversas.
Las membranas biológicas contienen una variedad de lípidos anfipáticos, incluyendo
los glicerofosfolípidos y los esfingolípidos. En algunos organismos, la función de los
triacilgliceroles (grasas y aceites) es de moléculas intracelulares de almacenamiento de
energía metabólica. También las grasas suministran aislamiento térmico y amortiguamiento
a los animales. Las ceras en las paredes celulares, en los exoesqueletos y en la
piel protegen a las superficies de algunos organismos. Algunos lípidos tienen funciones
muy especializadas. Por ejemplo, las hormonas esteroides regulan e integran una variedad
de actividades metabólicas en los animales, y los icosanoides participan en la regulación
de la presión sanguínea, en la temperatura corporal y en la contracción de los
músculos lisos en los mamíferos. Los gangliósidos y otros glicoesfingolípidos están en
la superficie celular y pueden participar en el reconocimiento celular.
*Ácidos grasos:
En varias especies se han identificado más de 100 ácidos grasos distintos. Los ácidos
grasos difieren entre sí en la longitud de sus colas de hidrocarburo, la cantidad de dobles
enlaces carbono-carbono, las posiciones de los dobles enlaces en las cadenas y la cantidad de ramificaciones. La mayor parte de los ácidos grasos tienen un pKa aproximado de 4.5 a 5.0, y por consiguiente están ionizados al pH fisiológico. Los ácidos grasos son una forma de
detergente, porque tienen una larga cola hidrofóbica y una cabeza polar .
Como es de esperar, la concentración de ácido graso libre en las células es muy baja, porque altas concentraciones de ácidos grasos libres podrían romper las membranas. La mayor parte de los ácidos grasos están formados por lípidos más complejos. Están unidos a otras moléculas mediante un enlace de éster en el grupo carboxilo terminal.
grasos difieren entre sí en la longitud de sus colas de hidrocarburo, la cantidad de dobles
enlaces carbono-carbono, las posiciones de los dobles enlaces en las cadenas y la cantidad de ramificaciones. La mayor parte de los ácidos grasos tienen un pKa aproximado de 4.5 a 5.0, y por consiguiente están ionizados al pH fisiológico. Los ácidos grasos son una forma de
detergente, porque tienen una larga cola hidrofóbica y una cabeza polar .
Como es de esperar, la concentración de ácido graso libre en las células es muy baja, porque altas concentraciones de ácidos grasos libres podrían romper las membranas. La mayor parte de los ácidos grasos están formados por lípidos más complejos. Están unidos a otras moléculas mediante un enlace de éster en el grupo carboxilo terminal.
Los ácidos grasos que no contienen dobles enlaces carbono-carbono se llaman saturados,
en tanto que los que tienen al menos un doble enlace carbono-carbono se clasifican
como no saturados o insaturados. Los ácidos grasos no saturados que sólo tienen un doble
enlace carbono-carbono se llaman monoinsaturados, en tanto que los que tienen dos
o más se denominan poliinsaturados. La configuración de los dobles enlaces en los
ácidos grasos no saturados es cis, en general. En la nomenclatura de IUPAC, se indican
las posiciones de los dobles enlaces mediante el símbolo Δn, en el que el superíndice n representa el átomo de carbono de menor número en cada par con doble enlace.
en tanto que los que tienen al menos un doble enlace carbono-carbono se clasifican
como no saturados o insaturados. Los ácidos grasos no saturados que sólo tienen un doble
enlace carbono-carbono se llaman monoinsaturados, en tanto que los que tienen dos
o más se denominan poliinsaturados. La configuración de los dobles enlaces en los
ácidos grasos no saturados es cis, en general. En la nomenclatura de IUPAC, se indican
las posiciones de los dobles enlaces mediante el símbolo Δn, en el que el superíndice n representa el átomo de carbono de menor número en cada par con doble enlace.
 |
| Aquí apreciamos las fuentes de los diferentes tipos de ácidos grasos. |
{kind=link}
 |
| Aquí la estructura de un ácido graso saturado, apreciamos que en esta estructura hay una ausencia total de insaturaciones (dobles enlaces) entre sus carbonos. |
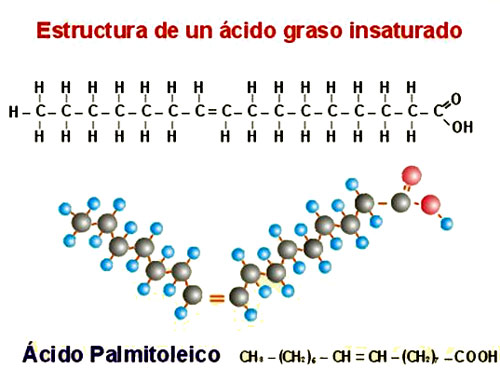 |
| Aquí también vemos un ácido graso, en este caso el ácido palmitoleico, apreciamos que en esta estructura si se ven dobles enlaces, lo que indica insaturaciones en sus estrucutura, pero son MONOINSATURADOS. |
 |
| Aquí vemos el omega 3 y omega 6 en orden descendente, este tipo de ácidos grasos, poseen muchas insaturaciones en su estructura por lo que se llaman ácidos grasos poliinsaturados, son un suplemento alimenticio encontrado en el hígado de ciertos pescados. |
*Triacilgliceroles:
La mayor parte de los lípidos en la dieta humana promedio son triacilgliceroles.Esos lípidos se descomponen en el intestino delgado por acción de las lipasas. Esas
enzimas se sintetizan como zimógenos en el páncreas, y son secretadas en el intestino
delgado, donde se activan. La lipasa pancreática cataliza la hidrólisis de los ésteres primarios (en el C-1 y C-3) de los triacilgliceroles, liberando los ácidos grasos y generando monoacilgliceroles. Como los lípidos no son solubles en el agua, la digestión de los lípidos se lleva a cabo en presencia de detergentes enérgicos, llamados sales biliares que son derivados anfipáticos del colesterol.Las micelas de las sales biliares
solubilizan los ácidos grasos y los monoacilgliceroles, de tal modo que se pueden
difundir y ser absorbidos por las células de la pared intestinal. Los lípidos se transportan
por el organismo en forma de complejos de lípido y proteína, llamados lipoproteínas.
En la mayor parte de las células, los triacilgliceroles coalescen en forma de gotitas
de grasa. Esas gotitas a veces se ven cerca de las mitocondrias en células que se basan
en ácidos grasos para obtener su energía metabólica. En los mamíferos, la mayor parte
de la grasa se almacena en el tejido adiposo, formado por células especializadas llamadas
adipocitos. Cada adipocito contiene una gota grande de grasa que forma casi todo el
volumen de la célula .
 |
| Vistas microscópicas de los adipocitos. |
*Glicerofosfolípidos:
Aunque los triacilgliceroles son los lípidos más abundantes en los mamíferos (por su
peso), no se encuentran en las membranas biológicas. Los lípidos más abundantes en la
mayor parte de las membranas son los glicerofosfolípidos (que también se llaman fosfoglicéridos);
como los triacilgliceroles tienen un soporte de glicerol. Los glicerofosfolípidos
más sencillos, los fosfatidatos, consisten en dos grupos acilo graso esterificados
en el C-1 y C-2 del 3-fosfato de glicerol (tabla 9.2). Nótese que hay tres grupos acilo
graso esterificados con la glicerina en los triacilgliceroles, en tanto que sólo hay dos
grupos acilo graso (R1 y R2) en los glicerofosfolípidos. La propiedad distintiva de los
grupos acilo (R1 y R2) de los glicerofosfolípidos es la presencia de un grupo fosfato en
el C-3 del soporte del glicerol. Las estructuras de los glicerofosfolípidos se pueden dibujar
como derivados del L-glicerol 3-fosfato (o 3-fosfato de L-glicerol), con el sustituyente
en el C-2 a la izquierda en una proyección de Fischer, como en la tabla 9.2. Para
simplificar, con frecuencia se mostrarán esos compuestos como estructuras no referidas
a su estereoquímica.
Los fosfatidatos están presentes en pequeñas cantidades como intermedios en la
biosíntesis o descomposición de glicerofosfolípidos más complejos. En la mayor parte
de los glicerofosfolípidos, el grupo fosfato está esterificado con glicerol y otro compuesto
que tenga un grupo — OH. Nótese que éstos son moléculas anfipáticas, con una cabeza polar y colas no polares. Por ejemplo, las membranas de los glóbulos rojos humanos contienen cuando menos 21 especies distintas fosfatidilcolina que difieren entre sí en las cadenas de acilo graso esterificadas en el C-1 y C-2 del respaldo de glicerina. En general, los glicerofosfolípidos tienen ácidos grasos saturados esterificados en el C-1 y ácidos grasos no saturados esterificados en el C-2.
peso), no se encuentran en las membranas biológicas. Los lípidos más abundantes en la
mayor parte de las membranas son los glicerofosfolípidos (que también se llaman fosfoglicéridos);
como los triacilgliceroles tienen un soporte de glicerol. Los glicerofosfolípidos
más sencillos, los fosfatidatos, consisten en dos grupos acilo graso esterificados
en el C-1 y C-2 del 3-fosfato de glicerol (tabla 9.2). Nótese que hay tres grupos acilo
graso esterificados con la glicerina en los triacilgliceroles, en tanto que sólo hay dos
grupos acilo graso (R1 y R2) en los glicerofosfolípidos. La propiedad distintiva de los
grupos acilo (R1 y R2) de los glicerofosfolípidos es la presencia de un grupo fosfato en
el C-3 del soporte del glicerol. Las estructuras de los glicerofosfolípidos se pueden dibujar
como derivados del L-glicerol 3-fosfato (o 3-fosfato de L-glicerol), con el sustituyente
en el C-2 a la izquierda en una proyección de Fischer, como en la tabla 9.2. Para
simplificar, con frecuencia se mostrarán esos compuestos como estructuras no referidas
a su estereoquímica.
Los fosfatidatos están presentes en pequeñas cantidades como intermedios en la
biosíntesis o descomposición de glicerofosfolípidos más complejos. En la mayor parte
de los glicerofosfolípidos, el grupo fosfato está esterificado con glicerol y otro compuesto
que tenga un grupo — OH. Nótese que éstos son moléculas anfipáticas, con una cabeza polar y colas no polares. Por ejemplo, las membranas de los glóbulos rojos humanos contienen cuando menos 21 especies distintas fosfatidilcolina que difieren entre sí en las cadenas de acilo graso esterificadas en el C-1 y C-2 del respaldo de glicerina. En general, los glicerofosfolípidos tienen ácidos grasos saturados esterificados en el C-1 y ácidos grasos no saturados esterificados en el C-2.

*Esfingolípidos:
y animales son los esfingolípidos. En los mamíferos tienen abundancia especial
en tejidos del sistema nervioso central. La mayor parte de las bacterias no tienen esfingolípidos.
El respaldo estructural de los esfingolípidos es la esfingosina (trans-4-esfingenina),
un alcohol no ramificado de C18, con un doble enlace trans entre el C-4 y C-5,
un grupo amino en el C-2 y grupos hidroxilo en el C-1 y C-3 (figura 9.10a). La ceramida
está formada por un grupo acilo graso unido al grupo amino del C-2 en la esfingosina, por un enlace de amida . Las ceramidas son los precursores metabólicos de todos los esfingolípidos. Las tres grandes familias de esfingolípidos son las esfingomielinas, los cerebrósidos y los gangliósidos. De ellos, sólo las esfingomielinas contienen fosfato, y se clasifican como fosfolípidos; los cerebrósidos y los gangliósidos contienen residuos de carbohidrato y se clasifican como glicoesfingolípidos.

unido a un enlace b-glicosídico al C-1 de una ceramida. Los galactocerebrósidos,
llamados también galactosilceramidas, tienen un solo residuo de b-D-galactosilo como
grupo de cabeza polar . Los galactocerebrósidos abundan en el tejido nervioso,
y forman casi el 15% de los lípidos en las vainas de mielina. Muchos otros tejidos en
los mamíferos contienen glucocerebrósidos, ceramidas con un grupo b-D-glucosilo
en la cabeza. En algunos glicoesfingolípidos, una cadena lineal hasta de tres residuos
más de monosacárido está unida a la mitad de galactosilo o glucosilo de un cerebrósido.
Los gangliósidos son glicoesfingolípidos más complejos, donde las cadenas de
oligosacárido que contienen ácido N-acetilneuramínico (NeuNAc) están unidas a una
ceramida. Se han caracterizado más de 60 variedades de gangliósidos. Su diversidad estructural
se debe a variaciones en la composición y la secuencia de los residuos de
azúcar. En todos los gangliósidos, la ceramida está unida a través de su C-1 a un residuo
b-glucosilo, que a su vez está enlazado a un residuo b-galactosilo.
Los gangliósidos están en las superficies celulares, con las dos cadenas de hidrocarburo
de la mitad de ceramida incrustadas en la membrana plasmática, y los oligosacáridos
en la superficie extracelular. Los gangliósidos y otros glicoesfingolípidos son
parte del repertorio en la superficie celular de diversas cadenas de oligosacárido, junto
con las glicoproteínas. En forma colectiva, esos marcadores proporcionan a las células
marcas superficiales distintivas que pueden servir en reconocimiento celular y comunicación de una célula a otra.
La composición de los glicoesfingolípidos de membrana puede cambiar en
forma notable durante el desarrollo de tumores malignos.
*Esteorides:Los esteroides son una tercera clase de lípidos que se encuentran en las membranas de los eucariotas, y muy rara vez en las bacterias. Los esteroides, junto con las vitaminas lipídicas y los terpenos, se clasifican como isoprenoides porque sus estructuras se relacionan con la molécula de isopreno, de cinco carbonos. Los esteroides contienen cuatro anillos fundidos, tres de seis carbonos identificados como A, B y C, y un anillo D de cinco carbonos. La estructura anular característica se deriva del escualeno. Los sustituyentes del sistema anular casi plano pueden apuntar hacia abajo (la configuración a) o hacia arriba (la configuración BETA). En la figura

*Lípidos no membranales: Hay muchos lípidos que no se encuentran en la membrana biológica de las células, entre estas varidades de lípidos están: ceras, eicosanoides y algunos isoprenoides.
-Ceras: Las ceras son ésteres no polares de ácidos grasos de cadena larga y alcoholes monohidroxílicos
de cadena larga. Por ejemplo, el palmitato de miricilo, uno de los principales
componentes de la cera de abejas, es el éster de palmitato y del alcohol
miricílico, de 30 carbonos . La hidrofobicidad del palmitato de miricilo hace
que la cera de abejas sea muy insoluble en agua, y su alto punto de fusión (debido a
las cadenas largas de hidrocarburo saturado) hace que esa cera sea dura y sólida a las
temperaturas peculiares de la intemperie. Las ceras están muy distribuidas en la naturaleza.
Proporcionan cubiertas protectoras impermeables a las hojas y frutos de ciertas
plantas, y en la piel, cuero, plumas y exoesqueletos de animales. La cera de oídos, llamada
también cerumen o cerilla (de cera, palabra latina que significa cera), es secretada
por las células que cubren el conducto auditivo. Lubrica el conducto y atrapa partículas
que podrían dañar al tímpano. La cera de oídos es una mezcla compleja, formada principalmente
por ácidos grasos de cadena larga, colesterol y ceramidas. También contiene
escualeno, triacilgliceroles y ceras verdaderas (10% del peso).



-Hay algunos lípidos que no están en membranas, que están relacionados con isoprenoides pero no son esteroides. Entre ellos están las vitaminas A,D,E y K.

¡Haz click aquí! y averigua más sobre la membrana celular y la acción de los lípidos en ella, son demasiado importantes para la vida
¡Observa el vídeo y descubre cualquier detalle no pulido o mencionado en este blog!
Lípidos en farmacia.
En farmacia se utilizan muchos lípidos como parte de las formulaciones que se dan tanto en la oficina de farmacia como en la industria farmacéutica en sí; aunque es estos son más utilizados para darle la forma farmacéutica al producto, más que como principios activos de la formulación, aunque existen sus excepciones.
*Aceite de Ricino:
Este aceite es obtenido por la extracción de la raíz de la planta del género Euphorbiaceae, Ricinus communis; es utilizado como un laxante natural y alivia el estreñimiento.
Su nombre quimico es ácido ricinoleico. Es un lípido monoinsaturado.
