Lípidos

Los lípidos más simples son los ácidos grasos, y tienen la fórmula general
R—COOH, donde R representa una cadena de hidrocarburo. Los ácidos grasos son
componentes de muchos tipos más complejos de lípidos, incluyendo los triglicéridos o
triacilgliceroles, los glicerofosfolípidos y los esfingolípidos. Los lípidos que contienen
grupos fosfato se llaman fosfolípidos y los que tienen grupos esfingosina y carbohidrato
a la vez se llaman glicoesfingolípidos. Los esteroides, las vitaminas lipídicas y los terpenos
se relacionan con la molécula de isopreno, de cinco carbonos, y por consiguiente se
llaman isoprenoides. El nombre terpenos se ha aplicado a todos los isoprenoides, pero
en general se restringe a los que existen en las plantas.
Los lípidos tienen funciones biológicas diversas, al igual que estructuras diversas.
Las membranas biológicas contienen una variedad de lípidos anfipáticos, incluyendo
los glicerofosfolípidos y los esfingolípidos. En algunos organismos, la función de los
triacilgliceroles (grasas y aceites) es de moléculas intracelulares de almacenamiento de
energía metabólica. También las grasas suministran aislamiento térmico y amortiguamiento
a los animales. Las ceras en las paredes celulares, en los exoesqueletos y en la
piel protegen a las superficies de algunos organismos. Algunos lípidos tienen funciones
muy especializadas. Por ejemplo, las hormonas esteroides regulan e integran una variedad
de actividades metabólicas en los animales, y los icosanoides participan en la regulación
de la presión sanguínea, en la temperatura corporal y en la contracción de los
músculos lisos en los mamíferos. Los gangliósidos y otros glicoesfingolípidos están en
la superficie celular y pueden participar en el reconocimiento celular.
R—COOH, donde R representa una cadena de hidrocarburo. Los ácidos grasos son
componentes de muchos tipos más complejos de lípidos, incluyendo los triglicéridos o
triacilgliceroles, los glicerofosfolípidos y los esfingolípidos. Los lípidos que contienen
grupos fosfato se llaman fosfolípidos y los que tienen grupos esfingosina y carbohidrato
a la vez se llaman glicoesfingolípidos. Los esteroides, las vitaminas lipídicas y los terpenos
se relacionan con la molécula de isopreno, de cinco carbonos, y por consiguiente se
llaman isoprenoides. El nombre terpenos se ha aplicado a todos los isoprenoides, pero
en general se restringe a los que existen en las plantas.
Los lípidos tienen funciones biológicas diversas, al igual que estructuras diversas.
Las membranas biológicas contienen una variedad de lípidos anfipáticos, incluyendo
los glicerofosfolípidos y los esfingolípidos. En algunos organismos, la función de los
triacilgliceroles (grasas y aceites) es de moléculas intracelulares de almacenamiento de
energía metabólica. También las grasas suministran aislamiento térmico y amortiguamiento
a los animales. Las ceras en las paredes celulares, en los exoesqueletos y en la
piel protegen a las superficies de algunos organismos. Algunos lípidos tienen funciones
muy especializadas. Por ejemplo, las hormonas esteroides regulan e integran una variedad
de actividades metabólicas en los animales, y los icosanoides participan en la regulación
de la presión sanguínea, en la temperatura corporal y en la contracción de los
músculos lisos en los mamíferos. Los gangliósidos y otros glicoesfingolípidos están en
la superficie celular y pueden participar en el reconocimiento celular.
*Ácidos grasos:
En varias especies se han identificado más de 100 ácidos grasos distintos. Los ácidos
grasos difieren entre sí en la longitud de sus colas de hidrocarburo, la cantidad de dobles
enlaces carbono-carbono, las posiciones de los dobles enlaces en las cadenas y la cantidad de ramificaciones. La mayor parte de los ácidos grasos tienen un pKa aproximado de 4.5 a 5.0, y por consiguiente están ionizados al pH fisiológico. Los ácidos grasos son una forma de
detergente, porque tienen una larga cola hidrofóbica y una cabeza polar .
Como es de esperar, la concentración de ácido graso libre en las células es muy baja, porque altas concentraciones de ácidos grasos libres podrían romper las membranas. La mayor parte de los ácidos grasos están formados por lípidos más complejos. Están unidos a otras moléculas mediante un enlace de éster en el grupo carboxilo terminal.
grasos difieren entre sí en la longitud de sus colas de hidrocarburo, la cantidad de dobles
enlaces carbono-carbono, las posiciones de los dobles enlaces en las cadenas y la cantidad de ramificaciones. La mayor parte de los ácidos grasos tienen un pKa aproximado de 4.5 a 5.0, y por consiguiente están ionizados al pH fisiológico. Los ácidos grasos son una forma de
detergente, porque tienen una larga cola hidrofóbica y una cabeza polar .
Como es de esperar, la concentración de ácido graso libre en las células es muy baja, porque altas concentraciones de ácidos grasos libres podrían romper las membranas. La mayor parte de los ácidos grasos están formados por lípidos más complejos. Están unidos a otras moléculas mediante un enlace de éster en el grupo carboxilo terminal.
Los ácidos grasos que no contienen dobles enlaces carbono-carbono se llaman saturados,
en tanto que los que tienen al menos un doble enlace carbono-carbono se clasifican
como no saturados o insaturados. Los ácidos grasos no saturados que sólo tienen un doble
enlace carbono-carbono se llaman monoinsaturados, en tanto que los que tienen dos
o más se denominan poliinsaturados. La configuración de los dobles enlaces en los
ácidos grasos no saturados es cis, en general. En la nomenclatura de IUPAC, se indican
las posiciones de los dobles enlaces mediante el símbolo Δn, en el que el superíndice n representa el átomo de carbono de menor número en cada par con doble enlace.
en tanto que los que tienen al menos un doble enlace carbono-carbono se clasifican
como no saturados o insaturados. Los ácidos grasos no saturados que sólo tienen un doble
enlace carbono-carbono se llaman monoinsaturados, en tanto que los que tienen dos
o más se denominan poliinsaturados. La configuración de los dobles enlaces en los
ácidos grasos no saturados es cis, en general. En la nomenclatura de IUPAC, se indican
las posiciones de los dobles enlaces mediante el símbolo Δn, en el que el superíndice n representa el átomo de carbono de menor número en cada par con doble enlace.
 |
| Aquí apreciamos las fuentes de los diferentes tipos de ácidos grasos. |
{kind=link}
 |
| Aquí la estructura de un ácido graso saturado, apreciamos que en esta estructura hay una ausencia total de insaturaciones (dobles enlaces) entre sus carbonos. |
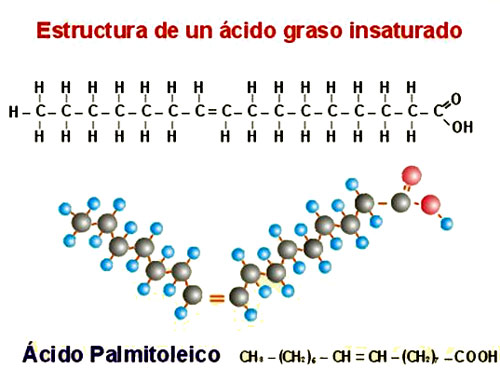 |
| Aquí también vemos un ácido graso, en este caso el ácido palmitoleico, apreciamos que en esta estructura si se ven dobles enlaces, lo que indica insaturaciones en sus estrucutura, pero son MONOINSATURADOS. |
 |
| Aquí vemos el omega 3 y omega 6 en orden descendente, este tipo de ácidos grasos, poseen muchas insaturaciones en su estructura por lo que se llaman ácidos grasos poliinsaturados, son un suplemento alimenticio encontrado en el hígado de ciertos pescados. |
*Triacilgliceroles:
La mayor parte de los lípidos en la dieta humana promedio son triacilgliceroles.Esos lípidos se descomponen en el intestino delgado por acción de las lipasas. Esas
enzimas se sintetizan como zimógenos en el páncreas, y son secretadas en el intestino
delgado, donde se activan. La lipasa pancreática cataliza la hidrólisis de los ésteres primarios (en el C-1 y C-3) de los triacilgliceroles, liberando los ácidos grasos y generando monoacilgliceroles. Como los lípidos no son solubles en el agua, la digestión de los lípidos se lleva a cabo en presencia de detergentes enérgicos, llamados sales biliares que son derivados anfipáticos del colesterol.Las micelas de las sales biliares
solubilizan los ácidos grasos y los monoacilgliceroles, de tal modo que se pueden
difundir y ser absorbidos por las células de la pared intestinal. Los lípidos se transportan
por el organismo en forma de complejos de lípido y proteína, llamados lipoproteínas.
En la mayor parte de las células, los triacilgliceroles coalescen en forma de gotitas
de grasa. Esas gotitas a veces se ven cerca de las mitocondrias en células que se basan
en ácidos grasos para obtener su energía metabólica. En los mamíferos, la mayor parte
de la grasa se almacena en el tejido adiposo, formado por células especializadas llamadas
adipocitos. Cada adipocito contiene una gota grande de grasa que forma casi todo el
volumen de la célula .
 |
| Vistas microscópicas de los adipocitos. |
*Glicerofosfolípidos:
Aunque los triacilgliceroles son los lípidos más abundantes en los mamíferos (por su
peso), no se encuentran en las membranas biológicas. Los lípidos más abundantes en la
mayor parte de las membranas son los glicerofosfolípidos (que también se llaman fosfoglicéridos);
como los triacilgliceroles tienen un soporte de glicerol. Los glicerofosfolípidos
más sencillos, los fosfatidatos, consisten en dos grupos acilo graso esterificados
en el C-1 y C-2 del 3-fosfato de glicerol (tabla 9.2). Nótese que hay tres grupos acilo
graso esterificados con la glicerina en los triacilgliceroles, en tanto que sólo hay dos
grupos acilo graso (R1 y R2) en los glicerofosfolípidos. La propiedad distintiva de los
grupos acilo (R1 y R2) de los glicerofosfolípidos es la presencia de un grupo fosfato en
el C-3 del soporte del glicerol. Las estructuras de los glicerofosfolípidos se pueden dibujar
como derivados del L-glicerol 3-fosfato (o 3-fosfato de L-glicerol), con el sustituyente
en el C-2 a la izquierda en una proyección de Fischer, como en la tabla 9.2. Para
simplificar, con frecuencia se mostrarán esos compuestos como estructuras no referidas
a su estereoquímica.
Los fosfatidatos están presentes en pequeñas cantidades como intermedios en la
biosíntesis o descomposición de glicerofosfolípidos más complejos. En la mayor parte
de los glicerofosfolípidos, el grupo fosfato está esterificado con glicerol y otro compuesto
que tenga un grupo — OH. Nótese que éstos son moléculas anfipáticas, con una cabeza polar y colas no polares. Por ejemplo, las membranas de los glóbulos rojos humanos contienen cuando menos 21 especies distintas fosfatidilcolina que difieren entre sí en las cadenas de acilo graso esterificadas en el C-1 y C-2 del respaldo de glicerina. En general, los glicerofosfolípidos tienen ácidos grasos saturados esterificados en el C-1 y ácidos grasos no saturados esterificados en el C-2.
peso), no se encuentran en las membranas biológicas. Los lípidos más abundantes en la
mayor parte de las membranas son los glicerofosfolípidos (que también se llaman fosfoglicéridos);
como los triacilgliceroles tienen un soporte de glicerol. Los glicerofosfolípidos
más sencillos, los fosfatidatos, consisten en dos grupos acilo graso esterificados
en el C-1 y C-2 del 3-fosfato de glicerol (tabla 9.2). Nótese que hay tres grupos acilo
graso esterificados con la glicerina en los triacilgliceroles, en tanto que sólo hay dos
grupos acilo graso (R1 y R2) en los glicerofosfolípidos. La propiedad distintiva de los
grupos acilo (R1 y R2) de los glicerofosfolípidos es la presencia de un grupo fosfato en
el C-3 del soporte del glicerol. Las estructuras de los glicerofosfolípidos se pueden dibujar
como derivados del L-glicerol 3-fosfato (o 3-fosfato de L-glicerol), con el sustituyente
en el C-2 a la izquierda en una proyección de Fischer, como en la tabla 9.2. Para
simplificar, con frecuencia se mostrarán esos compuestos como estructuras no referidas
a su estereoquímica.
Los fosfatidatos están presentes en pequeñas cantidades como intermedios en la
biosíntesis o descomposición de glicerofosfolípidos más complejos. En la mayor parte
de los glicerofosfolípidos, el grupo fosfato está esterificado con glicerol y otro compuesto
que tenga un grupo — OH. Nótese que éstos son moléculas anfipáticas, con una cabeza polar y colas no polares. Por ejemplo, las membranas de los glóbulos rojos humanos contienen cuando menos 21 especies distintas fosfatidilcolina que difieren entre sí en las cadenas de acilo graso esterificadas en el C-1 y C-2 del respaldo de glicerina. En general, los glicerofosfolípidos tienen ácidos grasos saturados esterificados en el C-1 y ácidos grasos no saturados esterificados en el C-2.

*Esfingolípidos:
y animales son los esfingolípidos. En los mamíferos tienen abundancia especial
en tejidos del sistema nervioso central. La mayor parte de las bacterias no tienen esfingolípidos.
El respaldo estructural de los esfingolípidos es la esfingosina (trans-4-esfingenina),
un alcohol no ramificado de C18, con un doble enlace trans entre el C-4 y C-5,
un grupo amino en el C-2 y grupos hidroxilo en el C-1 y C-3 (figura 9.10a). La ceramida
está formada por un grupo acilo graso unido al grupo amino del C-2 en la esfingosina, por un enlace de amida . Las ceramidas son los precursores metabólicos de todos los esfingolípidos. Las tres grandes familias de esfingolípidos son las esfingomielinas, los cerebrósidos y los gangliósidos. De ellos, sólo las esfingomielinas contienen fosfato, y se clasifican como fosfolípidos; los cerebrósidos y los gangliósidos contienen residuos de carbohidrato y se clasifican como glicoesfingolípidos.

unido a un enlace b-glicosídico al C-1 de una ceramida. Los galactocerebrósidos,
llamados también galactosilceramidas, tienen un solo residuo de b-D-galactosilo como
grupo de cabeza polar . Los galactocerebrósidos abundan en el tejido nervioso,
y forman casi el 15% de los lípidos en las vainas de mielina. Muchos otros tejidos en
los mamíferos contienen glucocerebrósidos, ceramidas con un grupo b-D-glucosilo
en la cabeza. En algunos glicoesfingolípidos, una cadena lineal hasta de tres residuos
más de monosacárido está unida a la mitad de galactosilo o glucosilo de un cerebrósido.
Los gangliósidos son glicoesfingolípidos más complejos, donde las cadenas de
oligosacárido que contienen ácido N-acetilneuramínico (NeuNAc) están unidas a una
ceramida. Se han caracterizado más de 60 variedades de gangliósidos. Su diversidad estructural
se debe a variaciones en la composición y la secuencia de los residuos de
azúcar. En todos los gangliósidos, la ceramida está unida a través de su C-1 a un residuo
b-glucosilo, que a su vez está enlazado a un residuo b-galactosilo.
Los gangliósidos están en las superficies celulares, con las dos cadenas de hidrocarburo
de la mitad de ceramida incrustadas en la membrana plasmática, y los oligosacáridos
en la superficie extracelular. Los gangliósidos y otros glicoesfingolípidos son
parte del repertorio en la superficie celular de diversas cadenas de oligosacárido, junto
con las glicoproteínas. En forma colectiva, esos marcadores proporcionan a las células
marcas superficiales distintivas que pueden servir en reconocimiento celular y comunicación de una célula a otra.
La composición de los glicoesfingolípidos de membrana puede cambiar en
forma notable durante el desarrollo de tumores malignos.
*Esteorides:Los esteroides son una tercera clase de lípidos que se encuentran en las membranas de los eucariotas, y muy rara vez en las bacterias. Los esteroides, junto con las vitaminas lipídicas y los terpenos, se clasifican como isoprenoides porque sus estructuras se relacionan con la molécula de isopreno, de cinco carbonos. Los esteroides contienen cuatro anillos fundidos, tres de seis carbonos identificados como A, B y C, y un anillo D de cinco carbonos. La estructura anular característica se deriva del escualeno. Los sustituyentes del sistema anular casi plano pueden apuntar hacia abajo (la configuración a) o hacia arriba (la configuración BETA). En la figura

*Lípidos no membranales: Hay muchos lípidos que no se encuentran en la membrana biológica de las células, entre estas varidades de lípidos están: ceras, eicosanoides y algunos isoprenoides.
-Ceras: Las ceras son ésteres no polares de ácidos grasos de cadena larga y alcoholes monohidroxílicos
de cadena larga. Por ejemplo, el palmitato de miricilo, uno de los principales
componentes de la cera de abejas, es el éster de palmitato y del alcohol
miricílico, de 30 carbonos . La hidrofobicidad del palmitato de miricilo hace
que la cera de abejas sea muy insoluble en agua, y su alto punto de fusión (debido a
las cadenas largas de hidrocarburo saturado) hace que esa cera sea dura y sólida a las
temperaturas peculiares de la intemperie. Las ceras están muy distribuidas en la naturaleza.
Proporcionan cubiertas protectoras impermeables a las hojas y frutos de ciertas
plantas, y en la piel, cuero, plumas y exoesqueletos de animales. La cera de oídos, llamada
también cerumen o cerilla (de cera, palabra latina que significa cera), es secretada
por las células que cubren el conducto auditivo. Lubrica el conducto y atrapa partículas
que podrían dañar al tímpano. La cera de oídos es una mezcla compleja, formada principalmente
por ácidos grasos de cadena larga, colesterol y ceramidas. También contiene
escualeno, triacilgliceroles y ceras verdaderas (10% del peso).



-Hay algunos lípidos que no están en membranas, que están relacionados con isoprenoides pero no son esteroides. Entre ellos están las vitaminas A,D,E y K.

¡Haz click aquí! y averigua más sobre la membrana celular y la acción de los lípidos en ella, son demasiado importantes para la vida
¡Observa el vídeo y descubre cualquier detalle no pulido o mencionado en este blog!
Lípidos en farmacia.
En farmacia se utilizan muchos lípidos como parte de las formulaciones que se dan tanto en la oficina de farmacia como en la industria farmacéutica en sí; aunque es estos son más utilizados para darle la forma farmacéutica al producto, más que como principios activos de la formulación, aunque existen sus excepciones.
*Aceite de Ricino:
Este aceite es obtenido por la extracción de la raíz de la planta del género Euphorbiaceae, Ricinus communis; es utilizado como un laxante natural y alivia el estreñimiento.
Su nombre quimico es ácido ricinoleico. Es un lípido monoinsaturado.

Ácidos Nucleicos


El
descubrimiento, en 1869, de la sustancia que resultó ser ácido
desoxirribonucleico (ADN) fue de Friedrich Miescher, joven médico suizo
que trabajaba en el laboratorio de Felix Hoppe-Seyler, químico fisiólogo
alemán. Miescher trató glóbulos blancos (contenidos en la pus de vendas
quirúrgicas desechadas) con ácido clorhídrico para obtener núcleos para
estudio. Cuando después se trataron los núcleos con ácido, se formó un
precipitado que contenía carbono, hidrógeno, oxígeno y un alto
porcentaje de fósforo. Miescher llamó “nucleína” al precipitado, porque
provenía de núcleos. Después, cuando se vio que era fuertemente ácido,
su nombre cambió a ácido nucleico. Aunque no lo supo, Miescher había
descubierto el ADN. Poco después, Hoppe-Seyler aisló una sustancia
parecida de las células de levadura. Hoy se sabe que esa sustancia es
ácido ribonucleico (ARN). Tanto ADN como ARN son polímeros de
nucleótidos, o polinucleótidos.

*Los nucleótidos:
 |
| Forma general para los nucleótidos de cada uno de los ácidos nucleicos. |
Los ácidos nucléicos como ya se ha mencionado, son polinucleótidos, los nucleótidos a su vez,están constituidos por un azúcar de cinco carbonos que ciclan en anillos furano, uno o más grupos fosfato y una base nitrogenada.
*Azúcares:
Las azúcares presentes en los nucléótidos, son: Ribosa y desoxirribisa. Estas,azúcares componentes de los nucleótidos que se encuentran en los ácidos nucleicosse muestran en la imagen inferior.Los dos azúcares aparecen como proyecciones de Haworth de la configuración b de las formas de anillo de furanosa . Es la configuración estable que existe en los nucleótidos y polinucleótidos. La conformación de la desoxirribosa predomina en el ADN de doble hebra.
 |
| Aquí podemos apreciar las conformaciones de Haworth para la Desoxirribosa (ADN) y de la Ribosa (ARN). |
*Bases nitrogenadas:
Las bases que se encuentran en los nucleótidos son derivados de pirimidina o de purina. La pirimidina tiene un solo anillo de cuatro átomos de carbono y dos de nitrógeno.La purina tiene un sistema de anillos fundidos de pirimidina y de imidazol. Los dos tipos
de bases son no saturados, con dobles enlaces conjugados. Esta propiedad hace que los
anillos sean planos, y también explica su capacidad de absorber la luz ultravioleta.
Las purinas y pirimidinas sustituidas son ubicuas en las células vivas, pero casi nunca
se encuentran las bases no sustituidas en los sistemas biológicos. Las principales pirimidinas que hay en los nucleótidos son uracilo (2,4-dioxopirimidina, U), timina
(2,4-dioxo-5-metilpirimidina, T) y citosina (2-oxo-4-aminopirimidina, C). Las principales purinas son adenina (6-aminopurina, A) y guanina (2-amino-6-oxopurina, G). Nótese que como la timina es una forma sustituida de uracilo, también se puede llamar 5-metiluracilo. La adenina, la guanina y la citosina están en ribonucleótidos y desoxirribonucleótidos. El uracilo se encuentra principalmente en ribonucleótidos y la timina en desoxirribonucleótidos.
Las purinas y las pirimidinas son bases débiles relativamente insolubles en agua al pH fisiológico. Sin embargo, dentro de las células la mayor parte de bases pirimidina y purina se encuentran como constituyentes de nucleótidos y polinucleótidos, compuestos que son muy hidrosolubles.
 |
| Es bueno, siempre recordar que el Uracilo está presente en la molécula del ARN; mientras que en el ADN está presente la Timina, una es homóloga de la otra pues ambas de conjugan con la Adenina, sólo que cada una en sus moléculas. |
dos formas tautómeras. La adenina y la citosina (que son amidinas cíclicas) pueden existir
en sus formas amino o imino, y la guanina, timina y uracilo (que son amidas cíclicas)
pueden existir en forma de lactama (ceto) o de lactima (enol) .
Las formas tautómeras de cada base existen en equilibrio, pero los tautómeros
amino y lactama son más estables, y en consecuencia predominan bajo las condiciones
que hay en el interior de la mayoría de las células. Los anillos permanecen no saturados
y planos en cada tautómero.
 |
| Aquí podemos apreciar as formás tautómeras de dos bases nitrogenadas. |
*Nucleósidos:
Los nombres de los nucleósidos se derivan de los de sus bases. El ribonucleósido que contiene adenina se llama adenosina (casi nunca se usa su nombre sistemático, 9-b-D ribofuranosiladenina); su contraparte desoxi se llama desoxiadenosina. De igual modo, los ribonucleósidos de guanina, citosina y uracilo son guanosina, citidina y uridina, respectivamente. Los desoxirribonucleósidos de guanina, citosina y timina son desoxiguanosina, desoxicitidina y desoxitimidina, respectivamente. Como casi nunca hay timidina en los ribonucleósidos, a la desoxitimidina se le llama con frecuencia sólo timidina. También por lo común se usan las abreviaturas de una sola letra para las bases de pirimidina y purina para designar a los ribonucleósidos: A, G, C y U para adenosina, guanosina, citidina y uridina, respectivamente. Los desoxirribonucleósidos se abrevian dA, dG, dC y dT, cuando es necesario distinguirlos de los ribonucleósidos. A veces está restringida la rotación de los enlaces glicosídicos en nucleósidos y nucleótidos. En los nucleósidos de purina, dos conformaciones, syn y anti, están en rápido equilibrio (figura 19.8, página siguiente). En los nucleósidos comunes de pirimidina predomina la conformación anti. En los ácidos nucleicos, que son polímeros de los nucleótidos, predominan las conformaciones anti.

*Nucleótidos:
Los nucleótidos son derivados fosforilados de los nucleósidos. Los ribonucleósidos
contienen tres grupos hidroxilo que se pueden fosforilar (2 , 3 y 5 ), y los desoxirribonucleósidos contienen dos de esos grupos hidroxilo (3 y 5 ). En los nucleótidos naturales,los grupos fosforilo suelen estar unidos al átomo de oxígeno del grupo 5 -hidroxilo. Por convención, siempre se supone que un nucleótido es un éster de 5 -fosfato, a menos que se indique otra cosa.

La doble helice de la molécula del Ácido Desoxirribonucleico.
La mayor parte de las moléculas de ADN consisten de dos hebras, de polinucleótidos.
Cada una de las bases en una hebra forma puentes de hidrógeno con una base de la hebra
opuesta . Los pares de bases más comunes están entre los tautómeros
lactama y amino de las bases. La guanina se aparea con citosina y la adenina con timina,
maximizando los puentes de hidrógeno entre sitios potenciales.

*Estabilización de la doble hélice por fuerzas débiles.
1. Interacciones de apilamiento Los pares de bases apilados forman contactos de
van der Waals. Aunque las fuerzas entre los pares de bases individuales apilados
son débiles, son aditivas, por lo que en las moléculas grandes de ADN los contactos
de van der Waals son una fuente importante de estabilidad.
2. Puentes de hidrógeno. Los puentes de hidrógeno entre pares de bases forman una
importante fuerza estabilizadora.
3. Efectos hidrofóbicos. Al sepultar los anillos hidrofóbicos de purina y pirimidina
en el interior de la doble hélice aumenta la estabilidad de la hélice.
4. Interacciones entre cargas. La repulsión electrostática de los grupos fosfato con
carga negativa en el esqueleto es una fuente potencial de inestabilidad de la hélice
de ADN. Sin embargo, la repulsión se minimiza por la presencia de cationes
como y proteínas catiónicas (que contienen abundancia de los residuos
básicos arginina y lisina).
¡Obten más información dando click aquí!
*Conformaciones del ADN.

Los estudios cristalográficos con rayos X de diversos oligodesoxirribonucleótidos
sintéticos, de secuencia conocida, indican que las moléculas dentro de la célula no
existen en una conformación B “pura”. En vez de ello, el ADN es una molécula dinámica
cuya conformación exacta depende hasta cierto grado de las secuencia de nucleótidos.
La conformación local también se afecta por dobleces en la molécula de ADN, y
de si está unida a una proteína. El resultado es que la cantidad de pares de bases por
vuelta en el B-ADN puede fluctuar de 10.2 a 10.6.
Además de varias formas de B-ADN, hay dos conformaciones muy diferentes del
ADN. Se forma A-ADN cuando se deshidrata el ADN, y se puede formar Z-ADN cuando
están presentes ciertas secuencias. (Las formas A y B de ADN fueron
descubiertas por Rosalind Franklin en 1952). El A-ADN está enrollado más apretadamente
que el B-ADN, y los surcos mayor y menor del A-ADN tienen ancho similar. Hay
unos 11 pb por vuelta en el A-ADN, y los pares de bases están desplazados unos 20° en
relación al eje mayor de la hélice. El Z-ADN difiere todavía más del B-ADN. En el ZADN
no hay surcos y la hélice es izquierda, no derecha. La conformación Z-ADN está en
regiones ricas en G/C. Los residuos de desoxiguanilato en el Z-ADN tienen distinta conformación de azúcares (3 -endo) y la base tiene la conformación sin. Tanto las conformaciones A-ADN como Z-ADN existen in vivo, pero se confinan a cortas regiones del ADN.
Superenrrollamiento de la molécula de ADN.

La mayor parte de las moléculas de ADN circular están superenrrolladas en las
células, pero hasta las moléculas lineales largas de ADN contienen regiones localmente
sobretorcidas. Los cromosomas bacterianos, en forma típica, tienen unos cinco superenrollamientos por 1 000 pares de bases de ADN. Según se verá en la sección 19.5, el ADN en los núcleos de las células eucarióticas también está sobretorcido. Todos los organismos tienen enzimas que pueden romper al ADN, destorcer o sobretorcer la doble hélice y volver a unir las hebras para alterar la topología.
células, pero hasta las moléculas lineales largas de ADN contienen regiones localmente
sobretorcidas. Los cromosomas bacterianos, en forma típica, tienen unos cinco superenrollamientos por 1 000 pares de bases de ADN. Según se verá en la sección 19.5, el ADN en los núcleos de las células eucarióticas también está sobretorcido. Todos los organismos tienen enzimas que pueden romper al ADN, destorcer o sobretorcer la doble hélice y volver a unir las hebras para alterar la topología.

Esas enzimas se llaman topoisomerasas, y se encargan de agregar y eliminar superenrollamientos. Estas notables enzimas rompen una o las dos hebras del ADN, desenvolviendo o envolviendo el ADN girando los extremos separados y los vuelven a unir, para crear (o eliminar) superenrollamientos.
1.ARN ribosómico (ARNr); moléculas que son parte integral de los ribosomas (ribonucleoproteínas intracelulares que son sitios de síntesis de proteínas). El ARN ribosómico es la clase más abundante de ácido ribonucleico, que forma 80% del ARN celular total.





Tipos de ARN presentes en las células
Las moléculas de ARN participan en varios procesos asociados a la expresión génica.
Esas moléculas se encuentran en copias múltiples y en varias formas distintas dentro de
una célula dada. Hay cuatro clases principales de ARN en todas las células vivas:
Esas moléculas se encuentran en copias múltiples y en varias formas distintas dentro de
una célula dada. Hay cuatro clases principales de ARN en todas las células vivas:
1.ARN ribosómico (ARNr); moléculas que son parte integral de los ribosomas (ribonucleoproteínas intracelulares que son sitios de síntesis de proteínas). El ARN ribosómico es la clase más abundante de ácido ribonucleico, que forma 80% del ARN celular total.

2. ARN de transferencia (ARNt); son moléculas que llevan a los aminoácidos activados a los ribosomas para su incorporación a las cadenas de péptidos en crecimiento durante la síntesis de proteínas. Las moléculas de ARNt sólo tienen de 73 a 95 residuos de nucleótidos de longitud. Forman un 15% del ARN celular total.


3. ARN mensajero(ARNm); moléculas que codifican las secuencias de aminoácidos en las proteínas. Son los “mensajeros” que llevan la información del ADN al complejo de traducción, donde se sintetizan las proteínas. En general, el ARNm sólo forma el 3% del ARN celular total. Estas moléculas son las menos estables de los ácidos ribonucleicos celulares.

4. ARN pequeño; moléculas presentes en todas las células. Algunas moléculas pequeñas de ARN tienen actividad catalítica o contribuyen a la actividad catalítica, asociadas a proteínas. Muchas de esas moléculas de ARN se relacionan con eventos de procesamiento que modifican al ARN después de que se ha sintetizado.

*La importancia de los ácidos nucleicos en Farmacia*.
La medicina va tomando un nuevo rumbo en cuanto a estrategias en el combate de enfermedades, así es como se presentó la ingienería genética, la cual basa su acción en los ácidos nucleicos. La nueva generación. En algunas instancias la acción de la ingienería genética se define como:
"La aplicación de diversos conocimientos sobre la biología a nivel molecular, en particular sobre el funcionamiento de los ácidos nucléicos (el ADN y los ARN), así como de las proteínas, ha conformado un área aplicada para profundizar la investigación y asimismo, el mejoramiento o desarrollo de nuevos productos, a partir de los procesos biológicos. Esta incluye las técnicas que permiten aislar y reproducir (clonar) genes; esto requiere una serie de enzimas (que ahora se consiguen a nivel comercial), que son capaces de cortar, modificar, pegar, sintetizar o degradar ácidos nucléicos. "Para poder cumplir con este objetivo operativo por parte de la ingienería genética se debe cumplir con los distintos requisitos:
- Disponer de un organismo donador de genes cuya función (es decir, la o las proteínas que codifican), se conozcan con suficiente profundidad. Estos genes se pueden aislar del genoma de origen o incluso, sintetizarse parcialmente en el laboratorio.
- Una forma de introducir este nuevo gen recombinante a células o tejidos de un organismo receptor, que serán transformados para adquirir una nueva función —o a veces suprimirla, como un carácter alergénico o cancerígeno— y, finalmente…
- Una forma de acondicionar o regenerar al organismo receptor final, para que se pueda utilizar de forma productiva con todas sus características más una (o dos o más).
- Insulina humana producida en bacterias:
Se necesitaba una fuente industrial para la insulina humana debido a
que las hasta ahora conseguidas, eran de origen animal, lo cual podía
generar respuestas inmunes. La insulina es el primer caso de proteína
producida por ingeniería genética aprobada para uso en humanos, desde
1982. La técnica de ingeniería
genética empleada consiste en extraer de células humanas el gen que
porta la información para fabricar insulina humana. Este gen (un
fragmento del material genético) se introduce dentro de bacterias que
son organismos fáciles de cultivar en el laboratorio.
Las bacterias que incorporaron el pequeño fragmento de ADN se denominan entonces "organismos genéticamente modificados" (OGM). Las bacterias que tienen el gen humano de la insulina se multiplican a un ritmo veloz y, a medida que lo hacen, producen grandes cantidades de insulina humana, entre otras sustancias. Entonces, la insulina humana se extrae de las bacterias, se purifica y se vende como medicamento. La sustancia obtenida por ingeniería genética, en este caso insulina humana, se denomina "insulina recombinante". - Terapia génica:La terapia génica es un campo emergente de la genética aplicada en la que se utilizan técnicas de ADN recombinante. En este caso, se emplean las propias moléculas de ADN recombinante con fines de tratamiento. La terapia génica consiste en la introducción de genes, creados mediante tecnología del ADN recombinante, en las células y los tejidos de los pacientes para tratar sus enfermedades. Los científicos están estudiando terapias génicas para tratar varias enfermedades humanas hereditarias en las que intervienen genes defectuosos. La idea es sustituirlos por genes funcionales nuevos.Desde el inicio del primer ensayo clínico en 1990, la investigación en terapia génica se ha extendido en gran medida, con un número cada vez mayor de ensayos en seres humanos. Este campo, aún en fases experimentales, centra sus esfuerzos en pacientes con enfermedades graves y potencialmente mortales que suelen contar con pocas opciones terapéuticas o en los que han fracasado todos los tratamientos disponibles.
Lo entendido en cuanto a material audiovisual sobre: Replicación del A.D.N.; Transcripción del A.D.N.;Traducción del A.D.N. a proteínas.
*Replicación del A.D.N.:
La replicación de ácido desoxirribonucleico, es semiconservativa debido a la presencia de una cadena parenteral que actúa como molde para la síntesis de nuevas cadenas complementarias.El primer paso se da con la ayuda de la helicasa, lo que consiste en desenrrollar la molécula del A.D.N. y romper los puentes de hidrógeno; las proteínas enlazantes de cadena sencilla evitan que las cadenas se vuelvan a unir, eso crea "burbujas de replicación" (segmentos donde las dos hebras del A.D.N., no están unidas) en diferentes partes de la molécula de A.D.N.; esto acelera la velocidad de la replicación.Para que se pueda dar la formación de una nueva cadena de A.D.N. del lado de la hebra conductora; una A.D.N. polimerasa es la encargada de la replicación en sentido de la horquilla de replicación, es decir, 5' hacia 3'. Pero sucede que la A.D.N. polimerasa solo puede elongar cadenas que ya hayan sido iniciadas y la enzima encargada de iniciar la cadena es la A.R.N. primasa, la cual agrega los primeros nucleótidos de A.R.N. cebador, los cuales posteriormente serán reemplazados por otros nucleótidos que serán de A.D.N., gracias a la labor de otra A.D.N. polimerasa. Esto sucede de manera continua en la cadena conductora de A.D.N. gracias a que la elongación sucede en orientación la horquilla de replicación mediante la cual es mucho más sencilla la labor de de replicación de esta cadena. Más específicamente, la adición de nucleótidos por parte de la polimerasa, se da cuando esta, trae un nucleótido trifosfato, el cual pierde 2 de sus grupos fosfatos, al romperse ese enlace, la energía que se produce es utilizada para la unión del grupo fosfato que permanece unido al carbono 5 del azúcar y éste se une al grupo OH.En la hebra rezagada, la polimerización se da de forma discontinua, debido a que este proceso en esta hebra, se hace en el sentido contrario al de la horquilla de replicación por lo cual debe esperar a que la hebra se desenrrolle. Por estos inconvenientes, se hace necesaria la labor de: la A.R.N. primasa, la ligasa y de otra polimerasa diferente a la aquella que realiza la adición de nucleótidos. La A.R.N. primasa, coloca los nucleótidos del A.R.N. cebador desde el cual inicia la polimerización de nucleótidos por parte de la de la A.D.N. polimerasa, luego igual que con la cadena conductora, otra A.D.N. polimerasa sustituye en A.R.N. cebador con nucleótidos de A.D.N. y los espacios dejado por la polimerización discontinua, son cubiertos por la labor de la ligasa y de esta manera se logra una replicación exacta de ambas cadenas.*Transcripción del A.D.N.
Es cuando el A.D.N. pasa a copiarse al A.R.N.Comienza con el ensamblaje de factores al inicio del gen. Uno de los que se ensamblan es la A.R.N. polimerasa; cuando la A.R.N. polimerasa es liberada esta se deplaza rápidamente leyendo el gen y mientras va desenrrollando el gen,copia una de las hebras del A.D.N.Lo nucleótidos que se utilizan para crear la copia en A.R.N., se emparejan con el A.D.N., nucleotido a nucleotido. Copian cada una de sus bases nitrogenadas menos la timina, la cual es sustituida por el uracilo.* Traducción del A.D.N. a proteínas.
La estructura del A.R.N. mensajero posee una cola poli-A y contiene codones que codifican aminoácidos específicos y al final de su estructura poseen una capa metilada.La labor del A.R.N. mensajero es llevar el mensaje del A.D.N., del núcleo al ribosoma para producri una proteína específica. Se dice que el ribosoma está compuesta por una pequeña y una parte grande y el proceso inicia cuando la parte pequeña se coloca en el punto inical de los codones.Otra molécula importante es la de A.R.N. de transferencia que contiene anticodones complementarios. Cuando el ribosoma va leyendo el A.R.N. mensajero,su codón está siendo traducido en aminoácidos.La parte grande del ribosoma se une y forma un punto A y B; cuando se forman estos sitios cada uno de los A.R.N. de transferencia pasa del sitio a B al sitiio A y el A.R.N. y la cadena de aminoácidos presentes pasa de un A.R.N. transferencia, a otro, esto se llama prolongación.Cuando el codón de paro actúa entra el sitio A del ribosoma, la proteína es liberada y disocia.
No hay comentarios:
Publicar un comentario